REPRODUCCION CELULAR
6. 1. CROMOSOMAS EUCARIOTAS:
El ADN, o ácido desoxirribonucleico, es el material que contiene la información hereditaria en los humanos y casi todos los demás organismos. Casi todas las células del cuerpo de una persona tienen el mismo ADN. La mayor parte del ADN se encuentra en el núcleo celular (o ADN nuclear), pero también se puede encontrar una pequeña cantidad de ADN en las mitocondrias (ADN mitocondrial o ADNmt). Las mitocondrias son estructuras dentro de las células que convierten la energía de los alimentos para que las células la puedan utilizar.
La
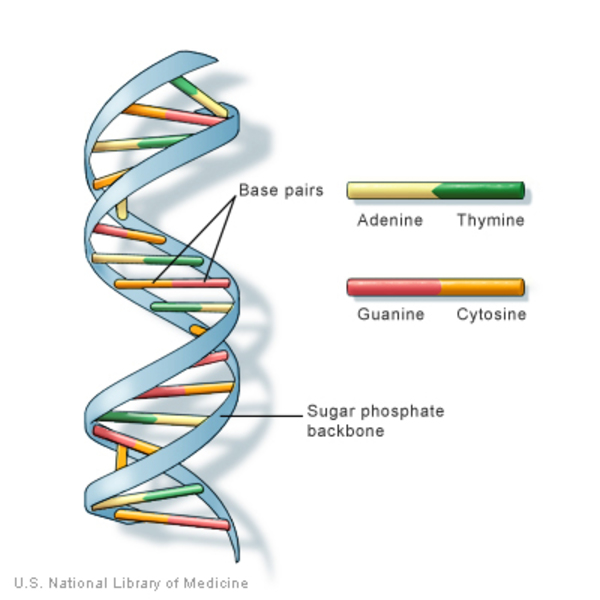
información en el ADN se almacena como un código compuesto por cuatro bases químicas, adenina (A), guanina (G), citosina (C) y timina (T). El ADN humano consta de unos 3 mil millones de bases, y más del 99 por ciento de esas bases son iguales en todas las personas. El orden o secuencia de estas bases determina la información disponible para construir y mantener un organismo, similar a la forma en que las letras del alfabeto aparecen en un cierto orden para formar palabras y oraciones.
Las bases de ADN se emparejan entre sí, adenina (A) con timina (T) y citosina (C) con guanina (G); para formar unidades llamadas pares de bases. Cada base también está unida a una molécula de azúcar y una molécula de fosfato. Juntos (una base, un azúcar y un fosfato) se llaman nucleótidos. Los nucleótidos están dispuestos en dos hebras largas que forman una espiral llamada doble hélice. La estructura de la doble hélice es algo parecido a una escalera, los pares de bases forman los peldaños de la escalera y las moléculas de azúcar y fosfato son sus pasamanos.
Una propiedad importante del ADN es que puede replicarse o hacer copias de sí mismo. Cada hebra de ADN en la doble hélice puede servir como patrón para duplicar la secuencia de bases. Esto es fundamental cuando las células se dividen, porque cada nueva célula necesita tener una copia exacta del ADN presente en la célula antigua.
El ADN es una doble hélice formada por pares de bases unidos a un esqueleto de azúcar-fosfato.
La metafase es una etapa de la división celular (mitosis o meiosis). Normalmente, los cromosomas individuales no son visibles en el núcleo celular. Sin embargo, durante la metafase en la mitosis o en la meiosis los cromosomas se condensan y se pueden distinguir cuando se alinean en el centro de la célula en división. Los cromosomas en la metafase (o metafásicos) se utilizan en la determinación del cariotipo que se realiza para buscar anormalidades cromosómicas.
Illustration
El estudio de la estructura externa de los cromosomas de cualquier especie eucariótica consiste en analizar la forma, tamaño y número de los cromosomas que posee. El mejor momento para llevar a cabo dicho estudio suele ser aquel en el que los cromosomas han alcanzado su máximo grado de contracción y tienen sus bordes perfectamente definidos. Dicho momento suele ser la metafase mitótica. El estudio de la estructura externa de los cromosomas culmina con la obtención del cariotipo.1
Los cromosomas se pueden estudiar en distintos momentos según la especie y dependiendo de los objetivos planteados. Algunas especies tienen cromosomas que se pueden observar con gran detalle en interfase, tal es el caso de Drosophila melanogaster, que posee cromosomas politénicos gigantes que se observan en las glándulas salivales de dicho insecto, y el de Chironomus tentans, otro díptero. El cariotipo se confecciona usualmente después de un apropiado pre-tratamiento y tinción de las células, para hacer más visibles los cromosomas individuales. Al diagrama simplificado de los cromosomas metafásicos del cariotipo se lo denomina idiograma, que se construye con el número genómico.
Para realizar el ordenamiento de los cromosomas tanto en cariotipos como idiogramas se debe tener en cuenta el tamaño cromosómico (ubicados de mayor a menor, con el brazo corto «bc» o «p» hacia arriba y el brazo largo «bl» o «q» hacia abajo); posición del centrómero (generalmente alineados) y presencia de constricciones secundarias y satélites.1
Constancia del número de cromosomas[editar]
| Especie | Número de cromosomas |
|---|---|
| Hormiga Myrmecia pilosula, macho | 1 |
| Hormiga Myrmecia pilosula, hembra | 2 |
| Mosca de la fruta (Drosophila melanogaster) | 8 |
| Centeno (Secale cereale) | 14 |
| Caracol (Helix) | 24 |
| Gato (Felis silvestris catus) | 38 |
| Cerdo (Sus scrofa) | 38 |
| Ratón (Mus musculus) | 40 |
| Trigo (Triticum aestivum) | 42 |
| Rata (Rattus rattus) | 42 |
| Conejo (Oryctolagus cuniculus) | 44 |
| Liebre (Lepus europaeus) | 46 |
| Humano (Homo sapiens sapiens) | 46 |
| Chimpancé (Pan troglodytes) | 48 |
| Patata, Papa (Solanum tuberosum) | 48 |
| Oveja (Ovis aries) | 54 |
| Vaca (Bos taurus) | 60 |
| Asno (Equus asinus) | 62 |
| Mula (Equus mulus) | 63 (estéril) |
| Caballo (Equus caballus) | 64 |
| Camello ( Camelus bactrianus) | 74 |
| Llama (Lama glama) | 74 |
| Perro (Canis lupus familiaris) | 78 |
| Gallina (Gallus gallus) | 78 |
| Paloma Columbia livia | 80 |
| Diamante mandarín (Taeniopygia guttata) | 7261 |
| Pez Carassius auratus | 94 |
| Equisetum arvense Equisetum arvense | 216 |
| Mariposa | 380 |
| Helecho Ophioglussum reticulatum | 1260 |
| Protozoario Aulacantha scolymantha | 1600 |
Usualmente las especies animales y vegetales tienen un número de cromosomas constante y determinado que constituyen su cariotipo (ley de la constancia numérica de los cromosomas), aunque existen especies con una alta variabilidad cariotípica, no solo en número sino en forma y tamaño de los cromosomas.

El número de cromosomas de una especie (o fase vital) diploide se identifica como 2n mientras que ese número en una especie (o fase vital) haploide se identifica con la letra n. En aquellas especies que presentan un número repetido de cromosomas superior a dos complementos se habla de poliploidía, representándose el múltiplo por delante de la letra n. Así: 3n indicaría un complemento cromosómico triploide, 4n un tetraploide, etc. Todas estas son situaciones de euploidía. Con la indicación x se quiere expresar el número básico de cromosomas de una especie que presenta individuos con diversos grados de ploidía o el de una línea filogenética a partir de la cual diversos taxones han alcanzado situaciones aneuploides variadas, siendo en este caso el número cromosómico una variación del número original con aumento o disminución del número básico, por pérdida, fusión o división de cromosomas (p. ej., n+1 o n-1). Un ejemplo de esta situación anormal la tenemos en los individuos de la especie humana que presentan el llamado síndrome de Down, situación de aneuploidía (2n=47) por la presencia de un ejemplar más de lo habitual del cromosoma 21 (trisomía).
El número de cromosomas 2n varía mucho de unas especies a otras y no existe relación entre el número de cromosomas y la complejidad de los mismos: existen especies vegetales con pocos cromosomas como Haplopappus gracilis (2n=4), Crepis capillaris (2n=6) y Secale cereale (2n=14), especies vegetales con bastantes cromosomas como Triticum aestivum (2n=42) y especies vegetales con muchos cromosomas como Ophioglossum petiolatum (n >500). En animales sucede algo semejante, hay especies con pocos cromosomas como la hormiga australiana Myrmecia pilosula cuyos machos tienen un cromosoma (2n=1) y las hembras dos cromosomas (2n=2), especies con bastantes cromosomas como la humana Homo sapiens (2n=46) y especies con muchos cromosomas como el lepidóptero Lysandra atlantica (2n=434-466). No existe ninguna relación entre el número de cromosomas 2n y la complejidad evolutiva, ni entre el número de cromosomas y la cantidad de ADN. Un ejemplo claro de esta situación es el de los ciervos del género Muntiacus en el que hay especies muy similares (denominadas especies gemelas) una con 2n=6 (M. muntjak) y otra con 2n=46 (M. reevesi).6364
Cromosomas sexuales[editar]
En muchos organismos, uno de los pares de los cromosomas homólogos es distinto al resto, realizando la determinación del sexo del individuo. A estos cromosomas se les llama cromosomas sexuales o heterocromosomas e incluso gonosomas, porque determinan el sexo.
- Sistema de determinación XY: es propio del ser humano y muchos otros animales. Las hembras, siendo XX, darán gametos iguales con cromosoma X, sexo homogamético y los machos, siendo XY, darán dos tipos de gametos, uno con el cromosoma X y otro con el cromosoma Y. La probabilidad de que en la fecundación, al unirse los gametos, resulte una combinación XX (hembra) o XY (macho) es aproximadamente del 50 %.
- Sistema de determinación ZW: en otras especies (p. ej. mariposas y aves) ocurre lo contrario, el sexo masculino es homogamético (ZZ) y el femenino heterogamético (ZW).
- Sistema de determinación XO: En otras especies (peces, insectos, anfibios, etc) que no tienen el cromosoma Y, determinándose el sexo por el número de cromosomas X, macho XO y hembra XX
.
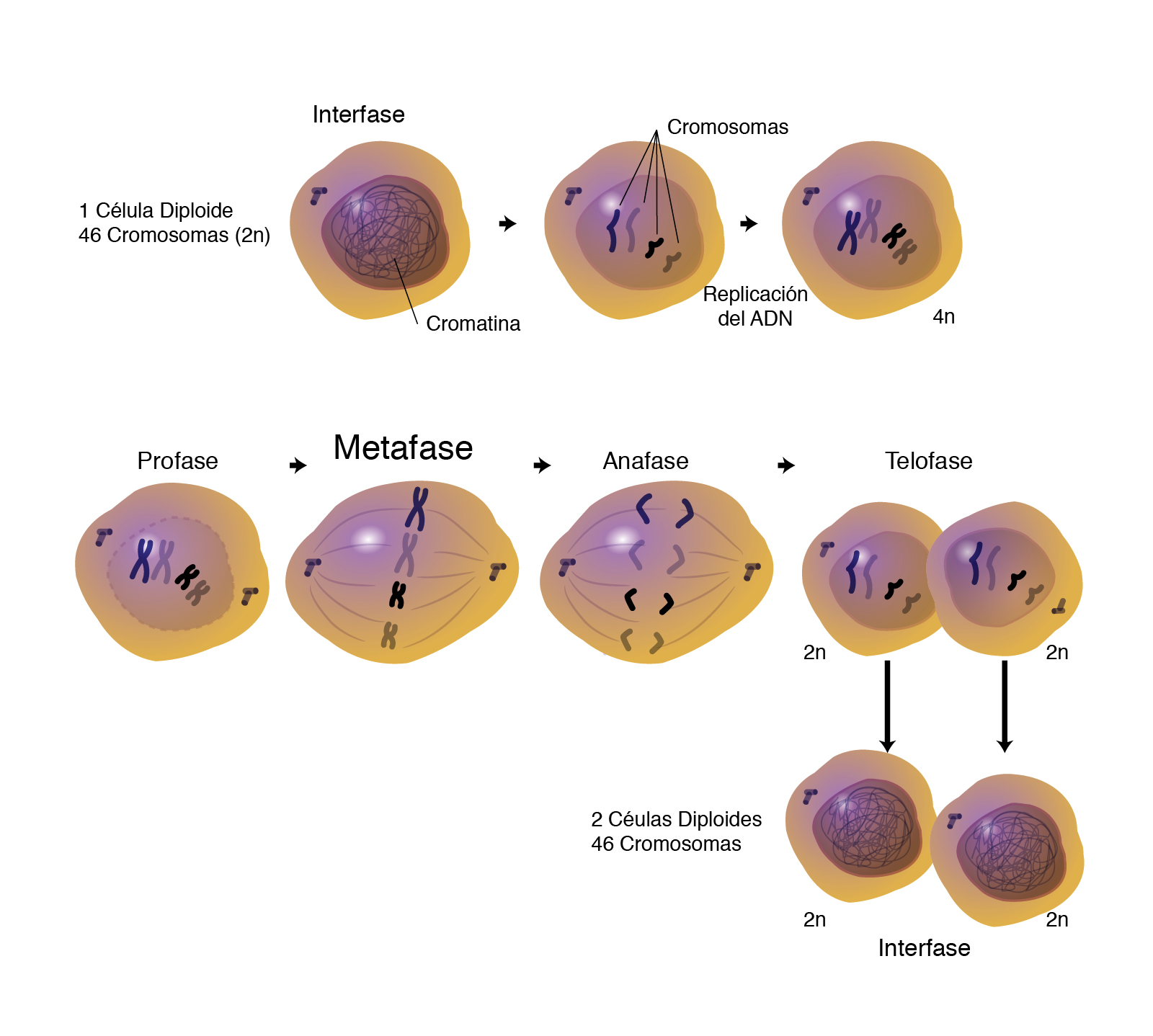
En biología, la mitosis es un proceso que ocurre en el núcleo de las células eucariotas y que precede inmediatamente a la división celular. Consiste en el reparto equitativo del material hereditario (ADN) característico.12 Este tipo de división ocurre en las células somáticas y normalmente concluye con la formación de dos núcleos (cariocinesis), seguido de otro proceso independiente de la mitosis que consiste en la separación del citoplasma (citocinesis), para formar dos células hijas.
La mitosis completa, que produce células genéticamente idénticas, es el fundamento del crecimiento, de la reparación tisular y de la reproducción asexual. La otra forma de división del material genético de un núcleo se denomina meiosis y es un proceso que, aunque comparte mecanismos con la mitosis, no debe confundirse con ella, ya que es propio de la división celular de los gametos. Produce células genéticamente distintas y, combinada con la fecundación, es el fundamento de la reproducción sexual y la variabilidad genética.
La mitosis es la división del núcleo celular en la que se conserva intacta la información genética contenida en los cromosomas, que pasa de esta manera sin modificaciones a las dos células hijas resultantes. La mitosis es igualmente un verdadero proceso de multiplicación celular que participa en el desarrollo, el crecimiento y la regeneración del organismo. Este proceso tiene lugar por medio de una serie de operaciones sucesivas que se desarrollan de una manera continua, pero para facilitar su estudio han sido separadas en varias etapas.


El resultado esencial de la mitosis es la continuidad de la información hereditaria de la célula madre en cada una de las dos células hijas. El genoma se compone de una determinada cantidad de genes organizados en cromosomas, hebras de ADN muy enrolladas que contienen la información genética vital para la célula y el organismo. Dado que cada célula debe contener completa la información genética propia de su especie, la célula madre debe hacer una copia de cada cromosoma antes de la mitosis, de forma que las dos células hijas reciban completa la información. Esto ocurre durante la fase S de la interfase, el período que alterna con la mitosis en el ciclo celular y en el que la célula entre otras cosas se prepara para dividirse.3
Tras la duplicación del ADN, cada cromosoma consistirá en dos copias idénticas de la misma hebra de ADN, llamadas cromátidas hermanas, unidas entre sí por una región del cromosoma llamada centrómero.4 Cada cromática hermana no se considera en esa situación un cromosoma en sí mismo, sino parte de un cromosoma que provisionalmente consta de dos cromáticas.
En animales y plantas, pero no siempre en hongos o protistas, la envoltura nuclear que separa el ADN del citoplasma se desintegra, desapareciendo la capa que separaba el contenido nuclear del citoplasma. Los cromosomas se ordenan en el plano ecuatorial de la célula, perpendicular a un eje definido por un huso acromático. Este es una estructura citoesquelética compleja, de forma ahusada, constituido por fibras que son filamentos de microtúbulos. Las fibras del huso dirigen el reparto de las cromátidas hermanas, una vez producida su separación, hacia los extremos del huso. Por convenio científico, a partir de este momento cada cromátida hermana sí se considera un cromosoma completo, y empezamos a hablar de cromosomas hermanos para referirnos a las estructuras idénticas que hasta ese momento llamábamos cromátidas. Como la célula se alarga, las fibras del huso «tiran» por el centrómero a los cromosomas hermanos dirigiéndolos cada uno a uno de los polos de la célula. En las mitosis más comunes, llamadas abiertas, la envoltura nuclear se deshace al principio de la mitosis y se forman dos envolturas nuevas sobre los dos grupos cromosómicos al acabar. En las mitosis cerradas, que ocurren por ejemplo en levaduras, todo el reparto ocurre dentro del núcleo, que finalmente se estrangula para formar dos núcleos separados.5
Se llama cariocinesis a la formación de los dos núcleos con que concluye habitualmente la mitosis. Es posible, y ocurre en ciertos casos, que el reparto mitótico se produzca sin cariocinesis (endomitosis) dando lugar a un núcleo con el material hereditario duplicado (doble número de cromosomas).
La mitosis se completa casi siempre con la llamada citocinesis o división del citoplasma. En las células animales la citocinesis se realiza por estrangulación: la célula se va estrechando por el centro hasta que al final se separa en dos. En las células de las plantas se realiza por tabicación, es decir, las células hijas “construyen” una nueva región de pared celular que dividirá la una de la otra dejando puentes de citoplasma (plasmodesmos). Al final, la célula madre se parte por la mitad, dando lugar a dos células hijas, cada una con una copia equivalente y completa del genoma original.
Cabe señalar que las células procariotas experimentan un proceso similar a la mitosis llamado fisión binaria. No se puede considerar que las células procariotas experimenten mitosis, dado que carecen de núcleo y únicamente tienen un cromosoma sin centrómero.6
Cariocinesis[editar]
La cariocinesis (del griego cario = núcleo y cinesis = movimiento), mitosis astral o mitosis anfiastral, es la división del núcleo celular. Consiste en la primera fase de la mitosis, que es el proceso por el cual el material genético de una célula madre se distribuye de manera idéntica entre dos células hijas.
En células animales poseen un organelo no membranoso llamado áster o centro celular, formado por un par de centriolos, que al dividirse en profase temprana, se dirigen hacia los polos opuestos de la célula, formando el aparato del huso mitótico, acrosómico o acromático.
Fases del ciclo celular[editar]
La división de las células eucariotas es parte de un ciclo vital continuo, el ciclo celular, en el que se distinguen dos períodos mayores, la interfase, durante la cual se produce la duplicación del ADN, y la mitosis, durante la cual se produce el reparto idéntico del material antes duplicado.
La mitosis es una fase relativamente corta en comparación con la duración de la interfase.
Interfase[editar]
Durante la interfase, la célula se encuentra en estado basal de funcionamiento. En dicha fase se lleva a cabo la replicación del ADN y la duplicación de los orgánulos para tener un duplicado de todo antes de dividirse. Es la etapa previa a la mitosis donde la célula se prepara para dividirse, en ésta, los centríolos y la cromatina se duplican, aparecen los cromosomas los cuales se observan dobles. El primer proceso clave para que se de la división celular es que todas las cadenas de ADN se dupliquen (replicación del ADN); esto se da inmediatamente antes de que comience la división, en un período del ciclo celular llamado interfase, que es aquel momento de la vida celular en que ésta no se está dividiendo. Tras la replicación tendremos dos juegos de cadenas de ADN, por lo que la mitosis consistirá en separar esas cadenas y llevarlas a las células hijas. Para conseguir esto se da otro proceso crucial que es la conversión de la cromatina en cromosomas.
La duración del ciclo celular en una célula típica es de 16 horas: 5 horas para G1, 7 horas para S, tres horas para G2 y 1 hora para la división. Este tiempo depende del tipo de célula que sea.3

Profase[editar]
Se produce en ella la condensación del material genético (ADN), para formar unas estructuras altamente organizadas, los cromosomas. Como el material genético se ha duplicado previamente durante la fase S de la Interfase, los cromosomas replicados están formados por dos cromátidas, unidas a través del centrómero por moléculas de cohesinas.
Uno de los hechos más tempranos de la profase en las células animales es la duplicación del centrosoma; los dos centrosomas hijos (cada uno con dos centriolos) migran entonces hacia extremos opuestos de la célula. Los centrosomas actúan como centros organizadores de unas estructuras fibrosas, los microtúbulos, controlando su formación mediante la polimerización de tubulina soluble.7 De esta forma, el huso de una célula mitótica tiene dos polos que emanan microtúbulos.
En la profase tardía desaparece el nucléolo y se desorganiza la envoltura nuclear.
Prometafase[editar]
La envoltura nuclear se ha disuelto, y los microtúbulos (verde) invaden el espacio nuclear. Los microtúbulos pueden anclar cromosomas (azul) a través de los cinetocoros (rojo) o interactuar con microtúbulos emanados por el polo opuesto. Esto se denomina mitosis abierta. Los hongos y algunos protistas, como las algas o las tricomonas, realizan una variación denominada mitosis cerrada, en la que el huso se forma dentro del núcleo o sus microtúbulos pueden penetrar a través de la envoltura nuclear intacta.89
Cada cromosoma ensambla dos cinetocoros hermanos sobre el centrómero, uno en cada cromátida. Un cinetocoro es una estructura proteica compleja a la que se anclan los microtúbulos.10 Aunque la estructura y la función del cinetocoro no se conoce completamente, contiene varios motores moleculares, entre otros componentes.11Cuando un microtúbulo se ancla a un cinetocoro, los motores se activan, utilizando energía de la hidrólisis del ATP para "ascender" por el microtúbulo hacia el centrosoma de origen. Esta actividad motora, acoplada con la polimerización/despolimerización de los microtúbulos, proporciona la fuerza de empuje necesaria para separar más adelante las dos cromátidas de los cromosomas.11
Cuando el huso crece hasta una longitud suficiente, los microtúbulos asociados a cinetocoros empiezan a buscar cinetocoros a los que anclarse. Otros microtúbulos no se asocian a cinetocoros, sino a otros microtúbulos originados en el centrosoma opuesto para formar el huso mitótico.12 La prometafase se considera a veces como parte de la profase.

Metafase[editar]
A medida que los microtúbulos encuentran y se anclan a los cinetocoros durante la prometafase, los centrómeros de los cromosomas se congregan en la "placa metafásica" o "plano ecuatorial", una línea imaginaria que es equidistante de los dos centrosomas que se encuentran en los 2 polos del huso.12 Este alineamiento equilibrado en la línea media del huso se debe a las fuerzas iguales y opuestas que se generan por los cinetocoros hermanos. El nombre "metafase" proviene del griego μετα que significa "después".
Dado que una separación cromosómica correcta requiere que cada cinetocoro esté asociado a un conjunto de microtúbulos (que forman las fibras cinetocóricas), los cinetocoros que no están anclados generan una señal para evitar la progresión prematura hacia la anafase antes de que todos los cromosomas estén correctamente anclados y alineados en la placa metafásica. Esta señal activa el checkpoint de mitosis.13

Anafase[editar]
Cuando todos los cromosomas están correctamente anclados a los microtúbulos del huso y alineados en la placa metafásica, la célula procede a entrar en anafase (del griego ανα que significa "arriba", "contra", "atrás" o "re-"). Es la fase crucial de la mitosis, porque en ella se realiza la distribución de las dos copias de la información genética original.
Entonces tienen lugar dos sucesos. Primero, las proteínas que mantenían unidas ambas cromátidas hermanas (las cohesinas), son cortadas, lo que permite la separación de las cromátidas. Estas cromátidas hermanas, que ahora son cromosomas hermanos diferentes, son separados por los microtúbulos anclados a sus cinetocoros al desensamblarse, dirigiéndose hacia los centrosomas respectivos.
A continuación, los microtúbulos no asociados a cinetocoros se alargan, empujando a los centrosomas (y al conjunto de cromosomas que tienen asociados) hacia los extremos opuestos de la célula. Este movimiento parece estar generado por el rápido ensamblaje de los microtúbulos.14
Estos dos estados se denominan a veces anafase temprana (A) y anafase tardía (B). La anafase temprana viene definida por la separación de cromátidas hermanas, mientras que la tardía por la elongación de los microtúbulos que produce la separación de los centrosomas. Al final de la anafase, la célula ha conseguido separar dos juegos idénticos de material genético en dos grupos definidos, cada uno alrededor de un centrosoma.

Telofase[editar]
La telofase (del griego τελος, que significa "finales") es la reversión de los procesos que tuvieron lugar durante la profase y prometafase. Durante la telofase, los microtúbulos no unidos a cinetocoros continúan alargándose, estirando aún más la célula. Los cromosomas hermanos se encuentran cada uno asociado a uno de los polos. La envoltura nuclear se reforma alrededor de ambos grupos cromosómicos, utilizando fragmentos de la envoltura nuclear de la célula original. Ambos juegos de cromosomas, ahora formando dos nuevos núcleos, se descondensan de nuevo en cromatina. La cariocinesis ha terminado, pero la división celular aún no está completa.
Si a continuación no se produce la citocinesis, entonces se originará una célula binucleada. La polinucleación en los tejidos de muchos organismos, es un proceso genéticamente programado de citodiferenciación y desarrollo.15
El siguiente paso es la citocinesis, generalmente aparece en secuencia inmediata al terminar la cariocinesis.

Citocinesis[editar]
La citocinesis es un proceso independiente, que se inicia simultáneamente a la telofase. Técnicamente no es parte de la mitosis, sino un proceso aparte, necesario para completar la división celular. En las células animales, se genera un surco de escisión (cleavage furrow) que contiene un anillo contráctil de actina en el lugar donde estuvo la placa metafásica, estrangulando el citoplasma y aislando así los dos nuevos núcleos en dos células hijas.16 Tanto en células animales como en plantas, la división celular está dirigida por vesículas derivadas del aparato de Golgi, que se mueven a lo largo de los microtúbulos hasta la zona ecuatorial de la célula.17 En plantas esta estructura coalesce en una placa celular en el centro del fragmoplasto y se desarrolla generando una pared celular que separa los dos núcleos. El fragmoplasto es una estructura de microtúbulos típica de plantas superiores, mientras que algunas algas utilizan un vector de microtúbulos denominado ficoplasto durante la citocinesis.18 Al final del proceso, cada célula hija tiene una copia completa del genoma de la célula original. El final de la citocinesis marca el final de la fase M.

Consecuencias de la mitosis[editar]
Mediante el proceso mitótico, el material genético se divide en dos núcleos idénticos, con lo que las dos células hijas que resultan si se produce la división del citoplasma (citocinesis) serán genéticamente idénticas. Por tanto, la mitosis es un proceso de división conservativo, ya que el material genético se mantiene de una generación celular a la siguiente. La mayor parte de la expresión génica se detiene durante la mitosis, pero mecanismos epigenéticos funcionan durante esta fase, para "recordar" los genes que estaban activos en mitosis y transmitirlos a las células hijas.19
Errores en la mitosis[editar]
Aunque los errores en la mitosis son muy poco frecuentes, este proceso puede fallar, especialmente durante las primeras divisiones celulares en el cigoto. Los errores mitóticos pueden ser especialmente peligrosos para el organismo, porque el descendiente futuro de la célula madre defectuosa mantendrá la misma anomalía.
Un cromosoma puede no separarse durante la anafase. Este fenómeno se denomina "no-disyunción". Si esto ocurre, una célula hija recibirá dos cromosomas hermanos y la otra se quedará sin ninguno. Esto da lugar a que una célula tenga tres cromosomas que codifiquen la misma información genética (dos hermanos y un homólogo), una condición conocida como trisomía, y la otra célula, que solamente tiene un cromosoma (el cromosoma homólogo), tendrá monosomía. Estas células se consideran aneuploides, y la aneuploidía puede causar inestabilidad genética, un hecho frecuente en cáncer.20
La mitosis es un proceso traumático. La célula pasa por cambios drásticos en su estructura, algunos orgánulos se desintegran y se reconstruyen en cuestión de horas, y los microtúbulos tiran constantemente de los cromosomas. Por tanto, en ocasiones los cromosomas pueden dañarse. Un brazo del cromosoma se puede romper y perder un fragmento, causando deleción. El fragmento puede incorporarse incorrectamente a otro cromosoma no homólogo, causando translocación. Se puede integrar de nuevo al cromosoma original, pero en una orientación inversa, causando inversión. O se puede tratar erróneamente como un cromosoma separado, causando duplicación cromosómica.
Una parte de estos errores pueden detectarse por alguno de los puntos de control existentes a través del ciclo celular, lo cual produce una parada en la progresión celular, dando tiempo a los mecanismos reparadores a corregir el error. Si esto no ocurre, el efecto de estas anormalidades genéticas dependerá de la naturaleza específica del error. Puede variar de una anomalía imperceptible, a carcinogénesis o a la muerte del organismo.
Endomitosis
Artículo principal: Endomitosis
La endomitosis es una variante de la mitosis sin división nuclear o celular, lo que da lugar a células con muchas copias del mismo cromosoma en el mismo núcleo. Este proceso también se denomina endoreduplicación, y las células resultantes endopoliploides.21 Un ejemplo de una célula que sufre endomitosis es el megacariocito.22
Los puntos de control celular son mecanismo que aseguran la fidelidad de la división celular en las células. Tales puntos de control verifican si los procesos en cada fase del ciclo celular han sido completados con precisión antes de progresar hacia la siguiente fase. Han sido identificados múltiples puntos de control, aunque algunos son mejor conocidos que otros.
En 1986, Temple y Raff describieron el ciclo celular como un reloj;1 si éste fuera el caso, cada una de las fases funcionaría de acuerdo a una especie de reloj interno, que determinaría cuánto tiempo debería durar. Sin embargo, actualmente el ciclo celular se describe como las piezas de un dominó que, al caer, hacen que la siguiente caiga también: igualmente, para que una fase del ciclo celular tenga lugar, la fase anterior tiene que haber finalizado correctamente. Los puntos de control aseguran que una fase haya finalizado antes de pasar a la siguiente. Por tanto, los checkpoints son mecanismos de control que refuerzan la dependencia durante el ciclo celular.2 Los sucesos del ciclo celular de la mayor parte de los organismos están organizados en rutas dependientes, en los cuales el inicio de los sucesos tardíos depende de que los sucesos iniciales hayan finalizado correctamente. La existencia de un mecanismo de control se evidencia cuando una droga, un mutante u otra condición libera una relación de dependencia en el ciclo celular: el suceso secundario en una ruta determinada tiene lugar aunque no se hayan cumplido los prerrequisitos necesarios. Por ejemplo, para que tenga lugar la mitosis es necesario que se haya completado la replicación del ADN, pero esta dependencia puede eliminarse mediante mutación de proteínas concretas, de forma que la mitosis puede ocurrir aunque el ADN no haya terminado de replicarse. Esto implica que la dependencia se debe a la existencia de un mecanismo de control (un checkpoint en el ciclo celular) y no a una característica intrínseca de los propios procesos.
Una función importante de muchos puntos de control consiste en evaluar los daños en el ADN, los cuales se detectan por mecanismos sensores. Cuando se localiza el daño, el punto de control envía una señal que detiene el ciclo celular hasta que se realiza la reparación o, cuando no es posible repararlo, marca la célula para su destrucción por apoptosis (mecanismo efector). Todos los checkpoints que valoran daños en el ADN parece que utilizan el mismo mecanismo sensor-señal-efector.

La mayor parte de las células de un organismo adulto están diferenciadas y no se dividen. Algunos tipos celulares están diferenciados de forma terminal y no pueden volver a proliferar a lo largo de toda la vida del individuo: es el caso de las células del músculo esquelético y cardíaco, los adipocitos y las neuronas. Otros tipos celulares, sin embargo, permanecen en estado quiescente (G0) pero pueden ser estimuladas para re-entrar en el ciclo celular, como los linfocitos T y B, y los fibroblastos. Para ello, antes de entrar en fase S deben superar un punto de control denominado "punto de restricción" que se encuentra al final de la fase G1. Esto se consigue mediante el incremento en los niveles de ciclinas D inducido por los factores de crecimiento. Por el contrario, las células que tienen capacidad de división (las células madre) deben cesar de dividirse, salir del ciclo celular y entrar en la fase fase G0 para comenzar el proceso de diferenciación y convertirse en células con una función especializada. Las únicas células que se dividen continuamente en el humano adulto son las células madre hematopoyéticas y las células epiteliales del intestino.
Los principales puntos de control que verifican la progresión a través del ciclo celular en eucariotas son los siguientes:
Punto de Restricción[editar]
Es el primer punto de control del ciclo celular, al final de la fase G1, justo antes de entrar en la fase S.3 La mayor parte de las células se paran en este momento y entran en un estado de reposo denominado G0. Las células eucarióticas normalmente se detienen en este punto de control si las condiciones ambientales son adversas (falta de nutrientes, por ejemplo). En células animales este punto de control se denomina "punto de restricción", mientras que en levaduras se denomina "punto de inicio" (start).
Este momento en G1 fue descrito por primera vez en 1974 por Arthur Pardee, quien lo denominó "punto de restricción" R.4 Pardee observó que las células que han pasado el punto R pueden progresar a través de la fase S independientemente de la presencia de mitógenos.5 Además, Pardee identificó que este checkpoint no funcionaba correctamente en líneas celulares cancerosas. Las líneas celulares cancerosas que se utilizaron en este estudio estaban infectadas con el virus de simio 40 (SV40).4 El descubrimiento de que las proteínas oncogénicas de los virus tumorales (como el antígeno grande T de SV40, E1A de adenovirus y E7 de HPV), desactivan el checkpoint de G1/S mediante su interacción inhibidora con el producto del gen de retinoblastoma,67 proporcionó datos fundamentales para la comprensión de los mecanismos moleculares que subyacen en este mecanismo de control.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Este punto de control está controlado fundamentalmente por la acción de los inhibidores de CDKs (CKI) denominados p16 (también INK4A) y p21Waf1.8 Estas proteínas inhiben CDK4/6 y aseguran que no pueda interaccionar con la ciclina D1 para continuar el avance en el ciclo celular. En condiciones de crecimiento inducido o expresión oncogénica de ciclina D, este checkpoint se supera porque la expresión aumentada de ciclina D permite su interacción con CDK4/6. Una vez que se forman los complejos activos CDK4/6-ciclina D, éstos fosforilan la proteína supresora de tumores retinoblastoma (Rb) y otras proteínas de la misma familia (p130 y p107), lo que libera la inhibición del factor de transcripción denominado E2F. Esto a su vez genera la expresión de moléculas requeridas para la transición G1/S, como las ciclinas E y A, la timidina kinasa, la timidilato sintetasa, la ADN polimerasa α o la dihidrofolato reductasa (DHFR), entre otras. Las ciclinas E y A interaccionan y activan la quinasa dependiente de ciclina 2 (CDK2), el principal activador de la transición G1/S, que impulsa el paso hacia la fase S.
Checkpoint de daños en el ADN[editar]
La variación de la secuencia del ADN en la línea germinal es esencial para mantener la variabilidad genética y asegurar la aparición de modificaciones que permitan una mejor adaptación al medio. Sin embargo, en la línea somática los cambios genéticos normalmente son nocivos, y las células han desarrollado estrictos mecanismos de seguridad para detectar y corregir las posibles alteraciones que haya podido sufrir el ADN. Una única alteración genómica (una mutación) que produzca una simple modificación en la cantidad producida de una proteína, o la sustitución de un único aminoácido en su secuencia, puede desencadenar una serie de variaciones que culminen con la generación de un tumor. La aparición de este tipo de errores están asociados con los fallos en los mecanismos que deberían haber detectado y corregido la lesión en el ADN que causó la mutación, o en último término, dirigido la célula afectada hacia un proceso de muerte celular. Por ello, los mecanismos que aseguran la integridad del ADN son fundamentales para el correcto funcionamiento celular.
Las modificaciones en la secuencia del ADN pueden generarse por modificaciones químicas espontáneas de sus componentes, por errores durante la replicación o por daños infligidos al ADN, debido a la presencia de agentes endógenos (generados por el propio metabolismo celular normal) o exógenos (procedentes del exterior). Entre éstos se encuentran la radiación ionizante (IR) y determinados productos químicos (véase Daño del ADN). El daño en el ADN inicia una respuesta que activa diferentes mecanismos de reparación que reconocen lesiones específicas en el ADN, que son reparadas en el momento para recuperar la secuencia original del ADN. Asimismo, el daño en el ADN provoca una parada en el ciclo celular, que conlleva la alteración de numerosos procesos fisiológicos, que a su vez implica síntesis, transporte y degradación de proteínas. La comprensión de cuál es la señal que emana de las lesiones en el ADN y cómo afecta a las numerosas rutas celulares implicadas se ha obtenido en gran parte estudiando la sofisticada respuesta a las roturas de doble hebra (DSBs, por Double Strand Breaks), que está conservada desde levaduras hasta humanos. Esta respuesta es una intricada red de señalización movilizada fundamentalmente por dos proteínas kinasas asociadas: ATM (Ataxia-telangiectasia Mutated, el gen mutado en esta enfermedad humana) y ATR (ATM- and Rad3- Related).910
Transición G1/S[editar]
Este checkpoint impide la preparación para la replicación del ADN hasta que se haya eliminado el daño detectado en el ADN. Uno de los eventos clave en este proceso es la activación inducida por el daño en el ADN de la molécula p53. La activación de p53 incluye su fosforilación, acetilación y sumolación, además de su estabilización y su translocación hacia el núcleo celular. La fosforilación de p53 perturba su interacción con la ligasa E3 de ubiquitina hMdm2, lo que impide la ubiquitinación y degradación de p53. La principal responsable de la fosforilación de p53 en diferentes aminoácidos es ATM, que además contribuye a la estabilización de p53 mediante la fosforilación de hMdm2 (que inhibe su interacción con p53) y del factor de transcripción E2F1 (que activa la trascripción de çl,19ARF, un inhibidor de hMdm2). Una vez estabilizada, p53 entra en el núcleo celular e induce la transcripción de varios genes, entre los cuales se incluyen el inhibidor de kinasas dependientes de ciclinas (CDKs) denominado p21Waf1. Esta molécula inhibe la formación de los complejos ciclina D-CDK4, lo que impide la transición G1/S. Simultáneamente, p53 induce la transcripción de genes implicados en la reparación del ADN.
Alternativamente, si el daño genómico es demasiado grande para que pueda ser reparado, ATM induce la activación de una serie de rutas celulares que culminan en la muerte celular.
Intra S[editar]
ATM juega también un papel importante en el control de la respuesta al daño al ADN inducido por IR durante la fase S. En este caso, ATM fosforila e induce hChk2, que a su vez fosforila Cdc25A. Esta es una proteína que defosforila tirosinas y que, en células no irradiadas, defosforila CDK2 para promover la transición de G1 a S. Cuando hChk2 está activado y fosforila Cdc25A, promueve la degradación de esta fosfatasa por el proteasoma, lo que impide la activación de los complejos ciclina E-CDK2, provocando así una parada en la iniciación y progresión de la replicación del ADN.
Además, la activación de ATM inducida por IR produce la fosforilación de la proteína BRCA1, lo cual produce la disociación de BRCA1 de los complejos en los que se encuentra. BRCA1 se asocia a muchas proteínas que están implicadas en la respuesta al daño al ADN y hay múltiples evidencias que sugieren que BRCA1 realiza funciones múltiples en este proceso a través de sus proteínas asociadas. Entre estas, se encuentran proteínas implicadas en modificaciones de la cromatina (SWI/SNF, HDAC1/2, p300/CBP), proteínas implicadas en la reparación del ADN (BACH1, BLM...) y proteínas implicadas en el control de la transcripción (GADD45, BARD1...).11 BRCA1 es el gen de susceptibilidad al cáncer de mama y ovario, mutado en el 50% de todos los casos familiares de estos tipos de cáncer.
Transición G2/M[editar]
El checkpoint de G2/M evita que entren en mitosis células que o bien han terminado la replicación de su ADN y han sido posteriormente expuestas a agentes que lo han dañado, o bien células que han traspasado el checkpoint intra-S con daño en el ADN sin reparar.
ATM también es importante para el funcionamiento de este checkpoint. Como en el caso anterior, ATM es necesario para la fosforilación de hChk2, y hChk2 es necesario para el mantenimiento del checkpoint de G2/M. hChk2 fosforila e inhibe la fosfatasa hCdc25C, que por tanto no puede defosforilar y activar los complejos ciclina B1-CDK1, que son necesarios para promover la entrada en mitosis.
Por otro lado, aunque ATM es la primera molécula en activarse en la respuesta de daño al ADN, y es la responsable de la respuesta rápida e inmediata, ATR se une en una fase tardía y mantiene el estado fosforilado de sustratos específicos. Esta importante redundancia añade una mayor complejidad a la respuesta. Sin embargo, además de su papel en la fase tardía de la respuesta, ATR también responde a daños al ADN que no activan ATM, como tratamientos de radiación UV, horquillas de replicación bloqueadas e hipoxia, defosforilando al menos algunos de los sustratos de ATM, como p53 y BRCA1.910
Checkpoint del ensamblaje del huso[editar]
Durante la mitosis, la célula organiza un huso mitótico con dos polos, al cual se anclan los cromosomas mediante unas estructuras proteicas denominadas cinetocoros. Los cromosomas constan de dos moléculas idénticas de ADN, las cromátidas hermanas, que permanecen unidas hasta la anafase a través de unas proteínas denominadas cohesinas. El mecanismo que detecta que se ha formado correctamente un huso mitótico, que todos los cromosomas están asociados a dicho huso de manera bipolar, y que todos ellos se encuentran alineados en la placa metafásica es el denominado checkpoint de mitosis, de anafase o también punto de control del ensamblaje del huso, abreviado SAC por sus siglas en inglés (Spindle Assembly Checkpoint).12
Si uno de los cromosomas, por alguna razón, se retrasa durante el proceso de alineamiento, esta maquinaria produce una detención temporal de la progresión en el ciclo celular : la célula se detiene en metafase, dando tiempo a los mecanismos de reparación a resolver el problema detectado. Si pasado un tiempo, el problema no se ha corregido, la célula será abocada a un proceso de muerte celular, un mecanismo de seguridad para evitar que se produzca una situación de aneuploidía, generalmente con consecuencias graves para el organismo.
Cuando todos los cromosomas se congregan correctamente en la placa metafásica, el checkpoint de mitosis se inactiva, de manera que se produce el corte de las cohesinas que mantenían unidas las cromátidas hermanas, disparándose de este modo la entrada en anafase. Finalmente, ambos juegos de cromátidas hermanas se organizan alrededor de los polos de las células hijas en telofase, y por tanto ambas células reciben un complemento cromosómico completo.
Comentarios
Publicar un comentario